


Panagiotis Tassis
Profesor Adjunto de Medicina y Reproducción Porcina, Clínica de Animales de Granja, Facultad de Veterinaria, Universidad Aristóteles de Tesalónica, Grecia.
Email: [email protected]
Las micotoxinas son metabolitos secundarios producidos por ciertas especies de hongos (géneros Aspergillus, Penicillium, Fusarium, Alternaria y Claviceps) que pueden encontrarse en los cereales de todo el mundo.
⇰ Son producidas antes (hongos como patógenos de las plantas) o después de la cosecha de los granos, así como durante el almacenamiento (hongos que crecen saprofíticamente).
 Los datos globales muestran que hasta el 80% de los cultivos para piensos y alimentos están contaminados con micotoxinas, y que la contaminación de los granos con más de una micotoxina es común1,2.
Los datos globales muestran que hasta el 80% de los cultivos para piensos y alimentos están contaminados con micotoxinas, y que la contaminación de los granos con más de una micotoxina es común1,2.Las aflatoxinas (AFs, principalmente AFB1), los alcaloides del cornezuelo, las fumonisinas (FBs, principalmente FB1), y los tricotecenos (por ejemplo, DON, toxina T-2, nivalenol) la ocratoxina A (OTA) y la zearalenona (ZEN) son consideradas importantes por sus efectos en la salud de los verracos3,4.
 Entre ellas, DON, ZEN y FB1 son las micotoxinas más frecuentes producidas por las especies de Fusarium2.
Entre ellas, DON, ZEN y FB1 son las micotoxinas más frecuentes producidas por las especies de Fusarium2.
Según la Comisión Europea5-7, los niveles máximos de contaminación recomendados para estas micotoxinas en los piensos para cerdos son:
- ⇰ 0,9 mg DON/kg de pienso (piensos complementarios y completos).
- ⇰ 0,1 mg ZEN/kg de pienso para lechones y cerdas jóvenes y 0,25 mg ZEN/kg de pienso para cerdas y cerdos de engorde, respectivamente.
- ⇰ 5 mg FBs/kg de pienso.
- ⇰ 0,25 mg T2 y HT2)/kg de pienso compuesto.
- ⇰ 0,02 mg de AFB1/kg de pienso.
- ⇰ 0,05 mg de OTA/kg de pienso.
La relevancia de las características del semen de los verracos en el éxito reproductivo y la mejora de los rasgos de importancia económica (por ejemplo, la tasa de crecimiento, la eficiencia de la conversión alimenticia) en las granjas farrow-to-finish es indiscutible8.
 Las alteraciones reproductivas están reconocidas como la restricción más costosa para la producción animal eficiente9. Las alteraciones reproductivas están reconocidas como la restricción más costosa para la producción animal eficiente9. |
Hoy en día, la demanda de rendimiento reproductivo intensivo con el uso de cerdas hiperprolíficas, solo puede satisfacerse con la utilización de semen de verraco de alta calidad que respalde este exigente ritmo reproductivo10.
 Aproximadamente, el 90% de la reproducción porcina se ha realizado con inseminación artificial en las últimas dos décadas, el 99% de la cual se ha llevado a cabo con el uso de semen líquido y prolongadores a temperatura ambiente8,10.
Aproximadamente, el 90% de la reproducción porcina se ha realizado con inseminación artificial en las últimas dos décadas, el 99% de la cual se ha llevado a cabo con el uso de semen líquido y prolongadores a temperatura ambiente8,10.Los resultados de estudios anteriores con humanos, roedores y animales domésticos, han aportado pruebas de los efectos deletéreos de varios contaminantes sobre los gametos masculinos en varias etapas de la espermatogénesis (es decir, en el testículo) durante el paso por el epidídimo, y en los espermatozoides maduros, después de la eyaculación y durante la capacitación11.
| La presente revisión hará hincapié en los efectos de las principales micotoxinas sobre las características del semen y la fertilidad de los verracos. |

Principales mecanismos de acción de las micotoxinas significativas para los cerdos
Las citadas micotoxinas significativas para el ganado porcino, tienen mecanismos de acción específicos y propiedades toxicocinéticas que se están investigando continuamente (revisado por12,13).
DEOXINIVALENOL
El DON inhibe predominantemente la elongación de proteínas14. Los síntomas clínicos previstos en los casos de toxicosis por DON incluyen:
 Reducción de la ingesta de pienso
Reducción de la ingesta de pienso- Vómitos y náuseas
- Hemorragia/necrosis
- Diarrea
También se han atribuido al DON efectos sobre la función intestinal y la inmunosupresión15.
FUMONISINAS
La FB1 puede inhibir la biosíntesis de los esfingolípidos a través de la inhibición de la ceramida sintasa16, lo que da lugar a una alteración del metabolismo de los esfingolípidos y una posterior acumulación de esfinganina17.
 La acumulación de esfinganina en el hígado y los riñones se ha asociado con la apoptosis y la mitosis de las células, dando lugar a fibrosis e hiperplasia nodular, respectivamente18,19. La acumulación de esfinganina en el hígado y los riñones se ha asociado con la apoptosis y la mitosis de las células, dando lugar a fibrosis e hiperplasia nodular, respectivamente18,19. |
Otros posibles efectos tóxicos de la FB1 son:
- Inmunosupresión celular y humoral20,21
- Hiporexia y disminución de la ganancia de peso22
- Alteración de la barrera intestinal23 al afectar a la función de las uniones estrechas, lo que significa que, junto con su capacidad para deteriorar la función inmunitaria, puede provocar un aumento de la susceptibilidad de los cerdos a los patógenos oportunistas24

ZEARALENONA
La ZEN se asemeja al 17β-estradiol y, junto con sus metabolitos (principalmente α- y β- zearalenol: α- y β- ZEL) son sustratos para los receptores de estrógeno. Es capaz de ejercer efectos estrogénicos, perjudicando el sistema reproductivo y la fertilidad de los cerdos25.
⇰ La toxicosis por ZEN se ha asociado predominantemente con trastornos reproductivos en cerdas y verracos, junto con efectos sobre el intestino y el sistema inmunitario de los cerdos4.
TOXINA T-2
La toxina T-2 inhibe el inicio de la síntesis de proteínas26, afectando también a la síntesis de ADN y ARN, lo que dificulta el funcionamiento normal de las células27.
Se considera que es agudamente tóxico y causa14,28,29:
- Emesis y diarrea
- Letargia
- Inmunosupresión
- Necrosis, inflamación serosahemorrágica necrótica-ulcerosa del tracto digestivo
- Leucopenia (apoptosis de leucocitos)
- Hemorragia, shock y muerte
- Irritación oral/dérmica
OCRATOXINA A
La OTA tiene un mecanismo de acción bastante complejo que aún no se ha explorado por completo. Posiblemente, la inhibición de la síntesis de proteínas y la producción de energía, la inducción del estrés oxidativo, la formación de aductos de ADN, así como la apoptosis/necrosis y la detención del ciclo celular están implicados en su acción tóxica30.
 La toxicidad renal es la expresión predominante de la potencia tóxica de la OTA, aunque también puede inducir efectos teratogénicos, embriotóxicos, genotóxicos, neurotóxicos, inmunosupresores y carcinogénicos31,32.
La toxicidad renal es la expresión predominante de la potencia tóxica de la OTA, aunque también puede inducir efectos teratogénicos, embriotóxicos, genotóxicos, neurotóxicos, inmunosupresores y carcinogénicos31,32.
AFLATOXINAS
Las aflatoxinas tienen varios efectos toxicológicos con diferentes mecanismos, la mayoría de los cuales aún no se han dilucidado por completo.
Los efectos mutagénicos de la AFB1 se atribuyen principalmente al metabolito intermedio AFB1-exo-8,9 epóxido (AFBO)33.
⇰ Como molécula altamente inestable, el AFBO reacciona con macromoléculas celulares, incluyendo ácidos nucleicos, proteínas y fosfolípidos, para inducir varias alteraciones genéticas, metabólicas, de señalización y de estructura celular34-36.
⇰ También se han sugerido efectos de la AFB1 sobre la función y la integridad de las células a través de la inducción del estrés oxidativo37-39.
⇰ La AFB1 se ha relacionado con la genotoxicidad, la inmunotoxicidad y la intoxicación aguda al actuar sobre el ADN genómico, otras macromoléculas funcionales y las células inmunocompetentes40.
Los efectos de las micotoxinas sobre las características del semen y capacidad de fecundación del verraco
Los efectos de las micotoxinas producidas por las especies de Fusarium (DON, ZEN, FBs y toxina T-2)
Sin duda, las micotoxinas suponen una amenaza importante para la salud y el rendimiento reproductivo de los cerdos.
DON y ZEN son extremadamente significativos en cuanto a su impacto en los cerdos y se ha demostrado que ambos inducen efectos perjudiciales en el sistema reproductivo de las cerdas y los verracos4.
 Aunque los estudios han descrito los efectos del DON y de la toxina T-2 en las cerdas multíparas y nulíparas, los ovarios, el desarrollo folicular, los ovocitos porcinos y las células de la granulosa, faltan estudios in vivo sobre sus efectos en los verracos41-44.
Aunque los estudios han descrito los efectos del DON y de la toxina T-2 en las cerdas multíparas y nulíparas, los ovarios, el desarrollo folicular, los ovocitos porcinos y las células de la granulosa, faltan estudios in vivo sobre sus efectos en los verracos41-44.EFFECTOS DEL DON, ZEN & TOXINA T-2 SOBRE LAS CÉLULAS DE LEYDIG
Un reciente estudio in vitro sobre los efectos de DON, ZEN y la toxina T-2 en las células de Leydig porcinas, demostró la evidencia de un aumento de la apoptosis celular, así como la alteración del potencial de la membrana de las mitocondrias y la distribución del ciclo celular45.
⇰ Además, ZEN y DON redujeron significativamente la secreción de testosterona y progesterona en las células de Leydig (posible inhibición de la conversión de progesterona en testosterona), mientras que la toxina T-2 solo redujo la secreción de testosterona.
Los resultados demostraron que la exposición a ZEN, DON y toxina T-2 puede provocar una toxicidad reproductiva que implica la inhibición de la esteroidogénesis y la proliferación celular, lo que contribuye a la apoptosis celular inducida por la lesión mitocondrial en las células de Leydig porcinas.
 La toxina T-2 fue la más citotóxica para las células de Leydig, seguida de DON y ZEN, aunque todas las micotoxinas probadas ejercieron un efecto citotóxico en las células de Leydig, al prolongar los ciclos celulares en la fase G1 e impedir la transición de la fase G1 a la S.
La toxina T-2 fue la más citotóxica para las células de Leydig, seguida de DON y ZEN, aunque todas las micotoxinas probadas ejercieron un efecto citotóxico en las células de Leydig, al prolongar los ciclos celulares en la fase G1 e impedir la transición de la fase G1 a la S.
| Cabe destacar que las células de Leydig son muy importantes para el mantenimiento del fenotipo masculino y la espermatogénesis. Entre sus principales funciones se encuentran la síntesis y secreción de hormonas esteroideas46, teniendo una mayor susceptibilidad a la contaminación por micotoxinas debido a su alto contenido en lípidos47. |
EFECTOS DE LA ZEN SOBRE LA FERTILIDAD DEL VERRACO Y CARACTERÍSTICAS DEL SEMEN
Estudios anteriores han demostrado los efectos de la ZEN sobre la fertilidad y las características del semen de los verracos.
La ZEN se ha asociado con alteraciones como la reducción de:
- Los niveles de testosterona en suero y de la libido (con la ingesta de 40 mg de ZEN/kg de pienso entre las 14 y 18 semanas de edad)48.
- Peso de los testículos (reducción del 30% del peso tras la ingestión de 0,5-0,6 mg de ZEN/kg de pienso durante 64 días)49.
- La espermatogénesis (9 mg de ZEN/ kg de pienso desde los 32 días de vida hasta los 145 o 312 días de vida)50.
⇰ Sin embargo, algunos experimentos de alimentación con verracos y jabalíes silvestres, mostraron la ausencia de efectos perjudiciales o una disminución transitoria de los parámetros de motilidad del semen de los verracos51,52.
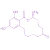
Los estudios in vitro han demostrado que ZEN y sus principales metabolitos α- y β-ZEL pueden disminuir la viabilidad de los espermatozoides y su motilidad progresiva:
- ZEN, α-ZEL, β-ZEL a 1 Χ 10-8 y 1 M o 2 Χ 10-7 y 20 μM durante 5, 16, 24 o 48 h53
- ZEN y α-ZEL a 125, 187,5 o 250 μM54
Además, la ZEN y α-ZEL pueden reducir la capacidad de los espermatozoides del verraco para unirse a la zona pelúcida (125,6, 188,4 y 251,2 μM ZEN, así como 124,8, 187,2 y 249,6 μM α-ZEL durante 1, 2, 3 o 4 h)55 y afectan a la integridad de la cromatina espermática (31,4, 62,8 y 94,2 μM ZEN, así como a-ZEL a niveles de 31,2, 62,4 y 93,6 μM durante 4 h)56.
Según Benzoni et al.53, ZEN, α- y β-ZEL influyen en una función particular de las células espermáticas.
⇰ Parece que el α-ZEL, que es el principal metabolito del ZEΝ en los cerdos, tenía mayor poder de desnaturalización de la estructura de la cromatina, mientras que la ZEΝ tenía una mayor potencia tóxica sobre la viabilidad del semen. El β-ZEL afectó exclusivamente a los parámetros de motilidad.
Por otra parte, se ha demostrado que, debido a su parecido estructural con el 17β-estradiol, la ZEN puede unirse a los receptores de estrógeno en varios tejidos, mientras que la α-ZEL se une a los receptores con mayor eficacia que la toxina madre57.
 Parece que los efectos perjudiciales sobre la fertilidad del verraco, podrían ser el resultado de la interacción entre ZEN o α- y β-ZEL y los receptores de estrógeno que se encuentran en los espermatozoides del verraco58.
Parece que los efectos perjudiciales sobre la fertilidad del verraco, podrían ser el resultado de la interacción entre ZEN o α- y β-ZEL y los receptores de estrógeno que se encuentran en los espermatozoides del verraco58.

EFECTOS DE ZEN & DON SOBRE EL SEMEN DE VERRACO
Nuestro grupo evaluó recientemente los efectos de DON y ZEN solos o en combinación, así como los efectos de sus formas modificadas, deepoxy-DON (DOM-1) y zearalenona hidrolizada (HZEN) en el semen de los verracos in vitro59,60.
⇰ Los resultados demostraron que las concentraciones de 50,6 μM de DON y 62,8 μM de ZEN indujeron efectos negativos significativos en las características del semen de los verracos in vitro, lo que sugiere un efecto negativo de estas micotoxinas en la fertilidad de los estos animales.
⇰ Los resultados de la evaluación del análisis de semen asistido por ordenador (CASA, computer-assisted semen analysis) (los puntos de tiempo de las observaciones del semen fueron 0, 1, 2, 3, 4 horas después de la exposición a las micotoxinas) sugirieron que un mayor número de espermatozoides inmóviles y no progresivos estaban presentes en todos los grupos expuestos a micotoxinas (es decir grupos DON, ZEN o DON + ZEN), independientemente del tiempo de incubación.
⇰ Se observó una reducción de los espermatozoides viables tras la exposición al ZEN, solo o en combinación con el DON (efecto del DON solo a la 4ª hora de observación). Los resultados de la prueba de hinchamiento hipoosmótico (HOST, Hypoosmotic swelling test) demostraron que ambas toxinas, solas o en combinación, pueden afectar negativamente a la integridad de la membrana de los espermatozoides. En cambio, se constató la ausencia de efectos sobre la integridad del ADN en todos los tipos de exposición59.
Cuando se aplicaron en combinación, DON y ZEN ejercieron un mayor efecto sobre muchos parámetros probados in vitro en comparación con la exposición única a DON o ZEN.

Los resultados de la comparación entre los efectos inducidos en el semen de los verracos tras la exposición in vitro a DON y ZEN en los niveles mencionados (50,6 μM DON y 62,8 μM ZEN, respectivamente) y la exposición a concentraciones equimolares de sus micotoxinas modificadas DOM-1 y HZEN aportaron más pruebas.
⇰ Se demostró que las formas modificadas no provocaban un deterioro de la motilidad, la morfología o la viabilidad de los espermatozoides, y que no inducían efectos negativos en la integridad de la membrana de los espermatozoides o de la cromatina nuclear.
 Estos resultados apoyan el hecho de que la conversión de DON en DOM-1 y de ZEN en HZEN representa un proceso de desintoxicación60.
Estos resultados apoyan el hecho de que la conversión de DON en DOM-1 y de ZEN en HZEN representa un proceso de desintoxicación60.EFECTOS DE FBs SOBRE EL SEMEN DE VERRACO
Las FBs también se han relacionado con efectos en el semen de los verracos.
- La FB1 a niveles de contaminación de 5, 10 o 15 mg/kg de pienso en cerdos después de 6 meses de ingestión de pienso contaminado (inicio de la alimentación a las 8-9 semanas de edad) puede asociarse con una reducción del vigor y del rendimiento reproductivo en los verracos.
- Se ha señalado que la FB1 en la dieta, por encima de 5 mg/kg, retrasa la consecución de la madurez sexual en cerdos en crecimiento9, reduciendo las reservas testiculares y epididimarias de esperma, así como la producción diaria de esperma y la calidad del semen en los verracos, con el potencial de perjudicar la fertilidad61,62.
Efectos de las micotoxinas producidas por especies de Aspergillus y Penicillium (AFBs y OTA)
EFECTOS DE LAS AFLATOXINAS SOBRE LA FERTILIDAD DEL VERRACO
Se ha demostrado la excreción de aflatoxinas en el semen de los verracos y sus posibles efectos adversos sobre su eficiencia reproductiva63.
En un estudio previo de 12 meses de duración, se evaluaron las concentraciones de AFB1 en el plasma seminal de 21 verracos de cuatro razas. Los resultados evidenciaron un aumento estacional de las concentraciones de AFB1 en el plasma seminal que estaba relacionado con los respectivos niveles de AFB1 en el pienso.
⇰ Además, los verracos con mayores niveles de AFB1 en el semen presentaron trastornos de la fertilidad, una menor tasa de supervivencia de los espermatozoides, así como una mayor proporción de espermatozoides anómalos.
EFECTOS DE LA OTA SOBRE LA FERTILIDAD DEL VERRACO
Los efectos de la OTA se evaluaron en un estudio con verracos (250 kg de peso) que recibieron 0,08 μg/kg de OTA por vía oral durante 6 semanas65. Los resultados indicaron una reducción de:
- La viabilidad espermática
- La motilidad inicial hacia delante
- Motilidad tras 24 horas de almacenamiento
En otro estudio in vivo66, se señaló que la OTA podría tener el potencial de afectar la producción espermática y la calidad seminal del verraco al provocar la reducción de la motilidad inicial y la longevidad de los espermatozoides.
⇰ Un estudio posterior planteó la hipótesis de que la OTA podría afectar a la espermatogénesis, ya que la duración del ciclo espermatogenético porcino es de 39 días y los resultados sugirieron una calidad espermática inferior aproximadamente 40 días después de comenzar el periodo de alimentación con toxinas.
⇰ La OTA es capaz de inhibir la síntesis de proteínas (interfiriendo con la fenilalanil- ARNt sintetasa que puede resultar en la modificación a nivel post-transcripcional de ARNm específicos) y este modo de acción específico podría estar también presente durante la espermatogénesis, resultando en una alteración que induciría una alteración de la estabilidad de la membrana del espermatozoide65,66.
Ewald y Heer67 encontraron una disminución de la producción espermática y calidad seminal del verraco tras la ingestión de OTA, como resultado de una mayor inestabilidad de la membrana espermática.
⇰ Según Zhang et al.68 se observó una disminución de la motilidad espermática en el verraco a un nivel de exposición de 100 μM de OTA durante 24 horas, mientras que las especies reactivas del oxígeno (ROS) aumentaron significativamente después de la exposición a 10 o 100 μM de OTA, y se observó un aumento de la expresión de los marcadores de apoptosis en el esperma expuesto a la OTA.
|
Según los resultados de este estudio, la exposición a la OTA afecta a la motilidad de los espermatozoides a través de las vías de señalización de la proteína quinasa activada (AMPK) y de PTEN (phosphatase and tensin homolog deleted on chromosome ten). Además, la generación de ROS en los espermatozoides puede activar al PTEN que, a su vez, conduce a la disminución de la AKT (proteína quinasa B) y la AMPK, y da lugar a una disminución de la motilidad de los espermatozoides69.
|
Efectos de transferencia en el embrión en desarrollo
Las interacciones durante la fecundación incluyen la entrega de componentes paternos desde el espermatozoide al ovocito. Estos componentes son fundamentales para70:
- ⇰ La activación del ovocito
- ⇰ La formación del cigoto
- ⇰ El desarrollo embrionario posterior
Por lo tanto, podría deducirse que los contaminantes ambientales (que entran a través de la alimentación en el cuerpo y son transportados al tracto reproductivo) que son capaces de modificar los componentes paternos, podrían afectar posteriormente al embrión en etapas posteriores del desarrollo.
 Los componentes paternos que podrían verse afectados incluyen el ADN, los marcadores epigenéticos, el ARN y las proteínas71.
Los componentes paternos que podrían verse afectados incluyen el ADN, los marcadores epigenéticos, el ARN y las proteínas71.
Los resultados, como los de los perfiles del transcriptoma de los blastocistos bovinos derivados de procedimientos de fecundación in vitro con espermatozoides tratados con AFB172, ponen de manifiesto la necesidad de seguir examinando el efecto de arrastre paterno en el embrión en desarrollo.
⇰ Los resultados de ese estudio demostraron la presencia de un gran número de genes transcritos de ARNm en los blastocistos desarrollados después de la fertilización con espermatozoides tratados, en comparación con los espermatozoides no tratados, por lo que se sugirió un posible efecto de arrastre de los espermatozoides al embrión en desarrollo.

CONCLUSIONES
Diversos estudios han demostrado los efectos in vitro e in vivo de varias micotoxinas en verracos y semen de verracos que podrían explicar la reducción de la fertilidad y el deterioro del rendimiento reproductivo en condiciones de campo.
Aunque la reprotoxicidad de las principales micotoxinas mencionadas se va revelando, es evidente que faltan pruebas sobre los efectos de varias otras micotoxinas, incluidas las emergentes, en la reproducción porcina. Además, no se han investigado suficientemente las posibles interacciones entre las micotoxinas sobre la espermatogénesis y la fertilidad del verraco.
⇰ Por lo tanto, es evidente la necesidad de realizar más estudios in vitro e in vivo sobre los efectos de las micotoxinas, solas o combinadas, haciendo hincapié en los mecanismos celulares y moleculares subyacentes que conducen a la alteración de las características del semen del verraco.
Además, teniendo en cuenta que la aparición y coocurrencia de micotoxinas es común en los piensos2, es fundamental aumentar la precaución para minimizar el riesgo de micotoxicosis en la granja para la salud y la espermatogénesis del verraco.

REFERENCIAS
1. Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R.Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2019.
2. Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 375.
3. Althouse, G.C. Applied Andrology in Swine. In Animal Andrology: Theories and Applications; Chenoweth, P.J., Lorton, S.P., Eds.; CABI: Oxfordshire, UK, 2014; pp. 404–417.
4. Ensley, S.M.; Radke, S.L. Mycotoxins in Grains and Feeds. In Disease of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 1055–1071.
5. European Commission (EC). Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. Off. J. Eur. Union. 2002, 32:1–22.
6. European Commission (EC). Recommendations on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off J Eur Union, 2013, 56, 12–15 L 91.
7. European Commission (EC).. Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union. 2006, 229:0007–0009.
8. Peltioniemi, O.A.; Kemp, B. Infertility in the Pig and the Control of Pig Herd Fertility. In Veterinary Reproduction and Obstetrics, 10th ed.; Noakes, E.D., Parkinson, T.J., England, G.C.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 581–592.
9. Gbore F.A. Growth performance and puberty attainment in growing pigs fed dietary fumonisin B1. Journal of Animal Physiology and Animal Nutrition, 2009a, 93: 761-767.
10. Rodriguez, A.L.; Van Soom, A.; Arsenakis, I.; Maes, D. Boar management and semen handling factors affect the quality of boar extended semen. Porc. Health Manag. 2017, 3, 15.
11. Komsky-Elbaz A, Kalo D, Roth Z. New evidence for deleterious effects of environmental contaminants on the male gamete. Anim Reprod Sci. 2021 Nov 6:106886.
12. Schelstraete W, Devreese M, Croubels S. Comparative toxicokinetics of Fusarium mycotoxins in pigs and humans. Food Chem Toxicol. 2020 Mar;137:111140.
13. Holanda DM, Kim SW. Mycotoxin Occurrence, Toxicity, and Detoxifying Agents in Pig Production with an Emphasis on Deoxynivalenol. Toxins (Basel). 2021 Feb 23;13(2):171.
14. Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Fund. Appl. Toxicol. 1984, 4, S124–132 2 PART 2.
15. Hooft JM, Bureau DP. Deoxynivalenol: Mechanisms of action and its effects on various terrestrial and aquatic species. Food Chem Toxicol. 2021
16. Wang E, Norred WP, Bacon CW, Riley RT, Merrill AH Jr. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with Fusarium moniliforme. J Biol Chem. 1991 Aug 5;266(22):14486-90.
17. Enongene, E.N.; Sharma, R.P.; Bhandari, N.; Miller, J.D.; Meredith, F.I.; Voss, K.A.; Riley, R.T. Persistence and reversibility of the elevation in free sphingoid bases induced by fumonisin inhibition of ceramide synthase. Toxicol. Sci. 2002, 67, 173–181.
18. Casteel, S.W.; Turk, J.R.; Cowart, R.P.; Rottinghaus, G.E. Chronic Toxicity of Fumonisin inWeanling Pigs. J. Veter Diagn. Investig. 1993, 5, 413–417.
19. Voss, K.A.; Plattner, R.D.; Riley, R.T.; Meredith, F.I.; Norred, W.P. In vivo effects of fumonisin B1-producing and fumonisin B1-nonproducing Fusarium moniliforme isolates are similar: Fumonisins B2 and B3 cause hepato- and nephrotoxicity in rats. Mycopathologia 1998, 141, 45–57.
20. Marin, D.E.; Taranu, I.; Pascale, F.; Lionide, A.; Burlacu, R.; Bailly, J.-D.; Oswald, I.P. Sex-related differences in the immune response of weanling piglets exposed to low doses of fumonisin extract. Br. J. Nutr. 2006, 95, 1185–1192.
21. Marin, D.E.; Gouze, M.-E.; Taranu, I.; Oswald, I.P. Fumonisin B1 alters cell cycle progression and interleukin-2 synthesis in swine peripheral blood mononuclear cells. Mol. Nutr. Food Res. 2007, 51, 1406–1412.
22. Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931.
23. Bouhet, S.; Hourcade, E.; Loiseau, N.; Fikry, A.; Martinez, S.; Roselli, M.; Galtier, P.; Mengheri, E.; Oswald, I.P. The mycotoxin fumonisin B1 alters the proliferation and the barrier function of porcine intestinal epithelial cells. Toxicol. Sci. 2004, 77, 165–171.
24. Oswald, I.P.; Desautels, C.; Laffitte, J.; Fournout, S.; Peres, S.Y.; Odin, M.; Le Bars, P.; Le Bars, J.; Fairbrother, J.M. Mycotoxin Fumonisin B1 Increases Intestinal Colonization by Pathogenic Escherichia coli in Pigs. Appl. Environ. Microbiol. 2003, 69, 5870–5874.
25. Zinedine, A., Soriano, J.M., Moltó, J.C., Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: an oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45 (1), 1–18.
26. Smith, K.E., Cannon, M., Cundliffe, E., 1975. Inhibition at the initiation level of eukaryotic protein synthesis by T-2 toxin. FEBS Lett. 50 (1), 2–6.
27. Sundstøl Eriksen, G., 2003. Metabolism and Toxicity of Trichothecenes. Doctoral Dissertation. Uppsala University, Uppsala, Sweden Department of Animal Nitrition and Management. Retrieved from. https://pub.epsilon.slu.se/287/.
28. Ueno, Y., 1977. Mode of action of trichothecenes. Pure Appl. Chem. 49 (4–6), 1737–1745.
29. Adhikari M, Negi B, Kaushik N, Adhikari A, Al-Khedhairy AA, Kaushik NK, Choi EH. T-2 mycotoxin: toxicological effects and decontamination strategies. Oncotarget. 2017, 8(20), 33933-33952.
30. Kőszegi T, Poór M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins (Basel). 2016 Apr 15;8(4):111.
31. Klarić MS, Rašić D, Peraica M. Deleterious effects of mycotoxin combinations involving ochratoxin A. Toxins 2013, 5(11):1965-87.
32. Yang S, Zhang H, De Saeger S, De Boevre M, Sun F, Zhang S, Cao X., Wang Z. In vitro and in vivo metabolism of ochratoxin A: a comparative study using ultra-performance liquid chromatography-quadrupole/time-of-flight hybrid mass spectrometry. Anal Bioanal Chem 2015, 407(13):3579-89.
33. Benkerroum, N. Retrospective and prospective look at aflatoxin research and development from a practical standpoint. Int. J. Environ. Res. Public Health 2019, 16, 3633.
34. Garner, R.C.;Wright, C.M. Binding of [14C]aflatoxin B1 to cellular macromolecules in the rat and hamster. Chem. Biol. Interact. 1975, 11, 123–131.
35. Zhuang, Z.; Huang, Y.; Yang, Y.; Wang, S. Identification of AFB1-interacting proteins and interactions between RPSA and AFB1. J. Hazard. Mater. 2016a, 301, 297–303.
36. Rushing, B.R.; Selim, M.I. Structure and oxidation of pyrrole adducts formed between aflatoxin B2a and biological amines. Chem. Res. Toxicol. 2017, 30, 1275–1285.
37. Bedard, L.L.; Massey, T.E. Aflatoxin B1-induced DNA damage and its repair. Cancer Lett. 2006, 241, 174–183.
38. Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2009, 38, 96–109.
39. Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438.
40. Benkerroum N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int J Environ Res Public Health. 2020 Jan 8;17(2):423.
41. Caloni, F.; Ranzenigo, G.; Cremonesi, F.; Spicer, L.J. Effects of a trichothecene, T-2 toxin, on proliferation and steroid production by porcine granulosa cells. Toxicon 2009, 54, 337–344.
42. Kanora A, Maes D. The role of mycotoxins in pig reproduction: A review. Vet Med-Czech. 2009; 54: 565- 576.
43. Cortinovis C, Pizzo F, Spicer LJ, Caloni F. Fusarium mycotoxins: effects on reproductive function in domestic animals–a review. Theriogenology. 2013 Oct 1;80(6):557-64.
44. Zhang Y, Han J, Zhu CC, Tang F, Cui XS, Kim NH, Sun SC. Exposure to HT-2 toxin causes oxidative stress induced apoptosis/autophagy in porcine oocytes. Sci Rep. 2016b; 6:1-8.
45. Sun, L.; Dai, J.; Xu, J.; Yang, J.; Zhang, D. Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro. Toxins 2022, 14, 113.
46. Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10.
47. Granum, C.; Anchersen, S.; Karlsson, C.; Berg, V.; Olsaker, I.; Verhaegen, S.; Ropstad, E. Steroidogenic differential effects in neonatal porcine Leydig cells exposed to persistent organic pollutants derived from cod liver oil. Reprod. Toxicol. 2015, 11, 130–139.
48. Berger, T.; Esbenshade, K.L.; Diekman, M.A.; Hoagland, T.; Tuite, J. Influence of prepubertal consumption of zearalenone on sexual development of boars. J. Anim. Sci. 1981, 53, 1559–1564.
49. Christensen, C.M.; Mirocha, C.J.; Nelson, G.H.; Quast, J.F. Effect of young swine of consumption of rations containing corn invaded by Fusarium roseum. Appl. Microbiol. 1972, 23, 202.
50. Young, L.G.; King, G.J. Low concentrations of zearalenone in diets of boars for a prolonged period of time. J. Anim. Sci. 1986, 63, 1197–1200.
51. Sutkeviciene N., B. Bakutis, A. Banys, B. Karveliene, A. Rutkauskas, J. Sabeckiene, Zilinskas H. The effect of the estrogenic mycotoxin zearalenone on boar reproductive potencial and the dynamic of aspartate aminotransferase and alanine aminotransferase levels in the boar blood serum. Veterinarija Ir Zootechnika (Vet. Med. Zoot.), 2009;46(68): 73-77.
52. Bielas W., W. Nizanski, J. Nicpon, J.E. Nicpon, A. Partyka, R. Mordak, M. Nowak, R. Ciaputa. Effect of zearalenone on circulating testosterone concentration, testicular and epididymal morphology and epididymal sperm characteristics in wild boars. Theriogenology 2017, 102 59-66.
53. Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol. 2008, 25, 461–467.
54. Tsakmakidis, I.A.; Lymberopoulos, A.G.; Alexopoulos, C.; Boscos, C.M.; Kyriakis, S.C. In vitro effect of zearalenone and alpha-zearalenol on boar sperm characteristics and acrosome reaction. Reprod. Domest. Anim. 2006, 41, 394–401.
55. Tsakmakidis, I.A.; Lymberopoulos, A.G.; Vainas, E.; Boscos, C.M.; Kyriakis, S.C.; Alexopoulos, C. Study on the in vitro effect of zearalenone and alpha-zearalenol on boar sperm-zona pellucida interaction by hemizona assay application. J. Appl. Toxicol. 2007, 27, 498–505.
56. Tsakmakidis, I.A.; Lymberopoulos, A.G.; Khalifa, T.A.; Boscos, C.M.; Saratsi, A.; Alexopoulos, C. Evaluation of zearalenone and alpha zearalenol toxicity on boar sperm DNA integrity. J. Appl. Toxicol. 2008, 28, 681–688.
57. Tiemann U, Dänicke S. In vivo and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non reproductive and reproductive organs in female pigs: a review. Food Addit Contam. 2007 Mar;24(3):306-14.
58. Rago V, Aquila S, Panza R, Carpino A. Cytochrome p450arom, androgen and estrogen receptors in pig sperm. Reprod Biol Endocrinol 2007;5:23, doi:10.1186/1477-7827-5-23.
59. Tassis, P.D.; Reisinger, N.; Nagl, V.; Tzika, E.; Schatzmayr, D.; Mittas, N.; Basioura, A.; Michos, I.; Tsakmakidis, I.A. Comparative Effects of Deoxynivalenol, Zearalenone and Its Modified Forms De-Epoxy-Deoxynivalenol and Hydrolyzed Zearalenone on Boar Semen In Vitro. Toxins 2022, 14, 497. https://doi.org/10.3390/toxins14070497
60. Tassis, P.D.; Tsakmakidis, I.A.; Nagl, V.; Reisinger, N.; Tzika, E.; Gruber-Dorninger, C.; Michos, I.; Mittas, N.; Basioura, A.; Schatzmayr, D. Individual and Combined In Vitro Effects of Deoxynivalenol and Zearalenone on Boar Semen. Toxins 2020, 12, 495.
61. Gbore F.A. Reproductive organ weights and semen quality of pubertal boars fed dietary fumonisin B1. Animal, 2009b, 3: 1133-1137.
62. Gbore F.A., Egbunike G.N.. Testicular and epididymal sperm reserves and sperm production of pubertal boars fed dietary fumonisin B1. Animal Reproduction Science, 2008, 105: 392-397.
63. Picha J, Cerovsky J, Pichova D (1986) Fluctuation in the concentration of sex steroids and aflatoxin B1 in the seminal plasma of boars and its relation to sperm production. Vet Med 1986, 31:347-357.
64. Kourousekos G. D., Theodosiadou E. Κ. (2018). Effects of aflatoxins on male reproductive system: A review. Journal of the Hellenic Veterinary Medical Society, 66(4), 201–210.
65. Biró K., Barna-Vetró I., Pécsi T., Szabó E., Winkler G., Fink-Gremmels J., Solti L. Evaluation of spermatological parameters in ochratoxin A—challenged boars. Theriogenology 2003, 60(2),199-207.
66. Solti L., Pécsi T., Barna-Vetró I., Szász F., Biró K., Szabó E., Analysis of serum and seminal plasma after feeding ochratoxin A with breeding boars. Anim. Reprod. Sci. 1999, 56:2, 123-132.
67. Ewald, C., Heer, A., 1989. The sperm quality of 4 boars during the assimilation of mycotoxin- containing feed (ochratoxin A) after changing the feed—a case study. Berl. Munch. Tierarztl. Wochenschr. 102, 261–266.
68. Zhang TY, Wu RY, Zhao Y, Xu CS, Zhang WD, Ge W, Liu J, Sun ZY, Zou SH, Shen W. Ochratoxin A exposure decreased sperm motility via the AMPK and PTEN signaling pathways. Toxicol Appl Pharmacol. 2018 Feb 1;340:49-57.
69. Zhao, Y., Zhang, W.D., Liu, X.Q., Zhang, P.F., Hao, Y.N., Li, L., Chen, L., Shen, W., Tang, X.F., Min, L.J., Meng, Q.S., Wang, S.K., Yi, B., Zhang, H.F., 2016. Hydrogen sulfide and/or ammonia reduces spermatozoa motility through AMPK/AKT-related pathways. Sci. Rep. 6 (37884).
70. Dogan, S., Vargovic, P., Oliveira, R., Belser, L.E., Kaya, A., Moura, A., Sutovsky, P., Parrish, J., Topper, E., Memili, E., 2015. Sperm protamine-status correlates to the fertility of breeding bulls. Biol. Reprod. 92, 92.
71. Castillo, J., Jodar, M., Oliva, R., 2018. The contribution of human sperm proteins to the development and epigenome of the preimplantation embryo. Hum. Reprod. Update 24, 535–555.
72. Komsky-Elbaz, A., Kalo, D., Roth, Z. Effect of aflatoxin B1 on bovine spermatozoa’s proteome and embryo’s transcriptome. Reproduction, 2020, 160.
 Prevención de micotoxicosis
Prevención de micotoxicosis