



Mariano Gorrachategui
Consultor en nutrición animal y presidente de la Comisión Técnica de CESFAC
Coocurrencia de micotoxinas
Sabemos que una micotoxina puede ser producida por varios tipos de hongos diferentes y que un mismo hongo toxigénico puede producir varias micotoxinas.
Además, hoy en día, el suministro de materias primas tiene orígenes muy diferentes y cereales, proteaginosas u oleaginosas crecen y se cosechan en condiciones climatológicas diversas que condicionan tanto el crecimiento fúngico como las toxinas producidas, además naturalmente de la evolución que existe en el almacenaje y que condiciona las toxinas que durante el mismo se pueden producir.
Con estos condicionantes no es difícil entender que la posibilidad de encontrar una única micotoxina como contaminante de materias primas o piensos es extremadamente baja. Por ello cada vez se usa más el término de “coocurrencia” de micotoxinas para referirnos a la presencia simultánea de dos o más micotoxinas en la misma matriz.

Existen muchas publicaciones en la literatura que ponen de manifiesto la coocurrencia de micotoxinas:
En Alemania, Goertz y col. (2010) observaron en maíz una contaminación con al menos 14 toxinas de Fusarium simultáneamente:
- ⇰ Deoxinivalenol (DON) y sus formas acetiladas
- ⇰ Zearalenona (ZEA)
- ⇰ Moniliformina (MON)
- ⇰ Beauvericina (BEA)
- ⇰ Nivalenol (NIV)
- ⇰ Eniantinas (ENNs)
- ⇰ Fumonisinas (FBs)
- ⇰ Toxina HT2
En un seguimiento global, Streit y col. (2013a) indicaron que, en piensos y materias primas, el 72% de las muestras contenían más de una micotoxina.
Los mismos autores (Streit y col., 2013b) observan en 83 muestras de piensos y materias primas una contaminación de entre 7 y 69 micotoxinas o metabolitos por muestra, detectando 169 compuestos diferentes.
Otros estudios han puesto de manifiesto la presencia simultánea de varias micotoxinas en muestras tomadas en países europeos (Almeida y col., 2011; Blajet-Kosicka y col., 2014; Driehuis y col., 2008; Labuda y col., 2005a, 2005b; Monbaliu y col., 2010) encontrándose en un elevado porcentaje de muestras la presencia simultánea de tricotecenos (DON, AcDON, T2, HT2) y FBs, con presencia de ZEA en bastantes casos.
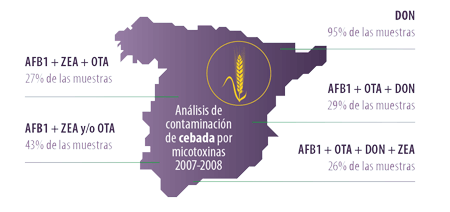
En España, cabe destacar el trabajo publicado por Ibáñez-Bea y col. (2012) sobre la presencia simultánea de AFB1, ZEA y OTA en muestras de cebada de las cosechas de 2007 y 2008, encontrando:
- ⇰ La presencia de las tres micotoxinas en el 27% de las muestras
- ⇰ La presencia de AFB1 con cualquiera de las otras dos en el 43%
Los mismos autores detectaron DON en el 95% de las muestras y dos micotoxinas de este grupo en el 43%.
Combinando todos los resultados de los estudios, concluyen que el 96% de las muestras contienen tres o más micotoxinas siendo las combinaciones más frecuentes:
- ⇰ AFB1, OTA y DON en el 29% de las muestras
- ⇰ AFB1, OTA, DON, y ZEA en el 26% de las muestras
Datos más recientes de Biomin (2017) en 1.378 muestras analizadas indican que el 94% de las muestras contenía más de 10 micotoxinas y metabolitos y que la media por muestra era de 28 micotoxinas.
En otro estudio, Raj et al. (2017), tras analizar la contaminación por micotoxinas de 113 muestras de maíz de Serbia y Bosnia y Hezergovina, determinaron que el 28% de las muestras estaba contaminada por más de una micotoxina. En otro estudio, los mismos autores encontraron un 76% de muestras contaminadas con una o más micotoxinas (Raj et al., 2019).
En forrajes también se han publicado datos recientes (Panasiuk y col., 2019).
Micotoxinas modificadas
Además de la coocurrencia, asumida de forma general, existe el fenómeno de la presencia de las hasta ahora denominadas “micotoxinas enmascaradas”.
Este término fue introducido por primera vez por Gareis y col. (1990) y surge porque en algunos casos de micotoxicosis, las observaciones clínicas en animales no se correspondían con el bajo contenido de micotoxinas determinado en el análisis del alimento.
Las micotoxinas enmascaradas se definían como “moléculas no detectables por análisis de rutina estándar”.
Años después, Berthiller y col. (2013) y, sobre todo, Rychlik y col. (2014) introdujeron los términos:
- ⇰ “Micotoxinas asociadas a las matrices” para aquellas micotoxinas asociadas a oligosacáridos o almidón, y que están físicamente atrapadas o están unidas por enlaces covalentes, como las Fumonisinas.
- ⇰ “Micotoxinas modificadas” que incluyen las:
- Micotoxinas “biológicamente” modificadas, por ejemplo, por conjugación, con compuestos polares, principalmente β-glucósido, sulfato o incluso glutatión, y donde se englobarían las denominadas “enmascaradas”.
- Micotoxinas “químicamente” modificadas, que se producen en los procesos térmicos o de otro tipo en la producción de piensos.
Micotoxinas emergentes
La evolución de los métodos analíticos basados en técnicas cromatográficas y espectrofotometría de masas permite detectar más compuestos de forma cada vez más fiable y en menor cantidad. Así, nos estamos encontrando con moléculas cuyos efectos tóxicos e interacciones son poco conocidas, siendo necesario conocerlas para producir alimentos seguros.
Nos referimos a las denominadas “micotoxinas emergentes”. Aunque este término no está claramente definido, nos referimos en general a:
- ⇰ Metabolitos de Fusarium como eniantinas (ENNs), beauvericina (BEA), moniliformina (MON), fusaproliferina (FP), ácido fusárico (FA), culmorina (CUL) y butenolida (BUT).
- ⇰ Metabolitos de Aspergillus como esterigmatocistina (STE) y emodina (EMO).
- ⇰ El metabolito de Penicillium, ácido micofenólico (MPA).
- ⇰ Metabolitos de Alternaria de las que hay más de 70 toxinas siendo, las más conocidas alternariol (AOH), monometil de alternariol éter (AME), altenueno (ALT), altertoxina (ATX) y ácido tenuazónico (TeA) (Gruber- Dorninger y col., 2017).
 Esta lista no es excluyente y, aunque actualmente estas micotoxinas no se analizan de forma rutinaria y no se contemplan en la legislación de alimentación animal, pueden tener alguna toxicidad e interaccionar con otras.
Esta lista no es excluyente y, aunque actualmente estas micotoxinas no se analizan de forma rutinaria y no se contemplan en la legislación de alimentación animal, pueden tener alguna toxicidad e interaccionar con otras.
¿Qué sabemos sobre la toxicidad de las micotoxinas emergentes?
ENNs & BEA
Las ENNs colonizan cereales y se pueden acumular en el grano. Sin embargo, aún no se las ha asociado a ninguna patología animal trascendente por vía natural (Marín García, 2010).
Según la EFSA (2014b), los efectos adversos para la salud de los animales por la exposición aguda a BEA y a la suma de ENNs en animales de granja y de compañía es poco probable. También es poco probable la aparición de efectos adversos por la exposición crónica en aves, sin que existan datos suficientes para valorarla en otras especies animales.
MON
En relación a la toxicidad de MON (EFSA, 2018), los datos disponibles para aves, cerdos y visones indican que la exposición a MON vía consumo de piensos presenta un riesgo bajo o insignificante para dichas especies según las prácticas actuales de alimentación.
Para el resto de especies, EFSA concluye que el riesgo es bajo o insignificante y que no se dispone de datos de toxicidad adecuados para poder caracterizar el peligro.
METABOLITOS DE ALTERNARIA
Los datos sobre la sensibilidad de los animales frente a las toxinas de Alternaria (EFSA, 2011) son muy limitados y no permiten la estimación de los niveles de tolerancia para toxinas individuales y mezclas de las mismas. Solo para las aves existen algunos datos de toxicidad para la evaluación del riesgo de estas toxinas.
La EFSA concluye que es poco probable que el AOH represente un riesgo en broilers, pero no pueden excluirse por completo los riesgos en la especie. La falta de datos toxicológicos no permite conclusión alguna para otras especies.
Interacciones entre micotoxinas
A veces se observan sintomatologías difíciles de explicar por la presencia de una única o varias micotoxinas o por la cantidad presente en pienso (Trenholm y col., 1983). Estos problemas se han relacionado con la interacción entre varias micotoxinas presentes, muchas de las cuales no se determinan analíticamente.
Esta teoría básica también explica el hecho reconocido de que los alimentos contaminados de forma natural tienen mayor toxicidad que los contaminados, de forma equivalente, con micotoxinas purificadas (Trenholm y col., 1994).
Klaasen y Eaton (1991) clasifican los efectos producidos por la interacción entre micotoxinas en:
- ⇰ Menos que aditivos
- ⇰ Aditivos
- ⇰ Sinérgicos
- ⇰ Potenciados
- ⇰ Antagonistas
Efectos sinérgicos de las micotoxinas
En general, sabemos o intuimos que en la mayoría de los casos existen efectos aditivos o sinérgicos (Speijers y Speijers, 2004; Pedrosa, 2010). Muchos autores han puesto de manifiesto esta aditividad, sinergia o potenciación:

Grenier y col. (2011) demostraron en maíz que los tumores hepáticos que inicia la presencia de AFB1 se ven potenciados por la presencia de FB1.
Además, tras revisar 112 publicaciones sobre las interacciones toxicológicas de las micotoxinas, Grenier y Oswald (2011) encontraron sinergias o aditividad en lo que se refiere al rendimiento en la mayoría de los estudios publicados. Sin embargo, en relación a otros parámetros, especialmente los bioquímicos, la respuesta es más variable yendo desde efectos sinérgicos hasta antagónicos para la misma combinación de toxinas.
Stoev y col. (2010) llegaron a una conclusión similar al estudiar nefropatías en pollos y cerdos que no se podían explicar solo por el contenido en OTA, por debajo del límite recomendado en la UE, explicándose por la presencia simultánea de OTA, FB1 y ácido penicílico (PCA).
Efectos antagónicos de las micotoxinas
Aunque la mayoría de los resultados muestran efectos aditivos o sinérgicos entre micotoxinas, es interesante indicar que también se observan efectos antagónicos (Koshinsky y Khachatourians, 1992; Bernhoft y col., 2004).

Así, se han observado antagonismos entre:
- ⇰ DON y FB1 (Ficheux y col., 2012; Wam y col., 2013a)
- ⇰ DON y ZEA (Bensassi y col., 2014; Wam y col., 2013)
- ⇰ DON y T2 (Thuvander y col., 1999; Ruíz y col., 2011)
- ⇰ DON y DAS (Thuvander y col., 1999)
- ⇰ NIV y DON o FB1 (Wam y col., 2013)
- ⇰ NIV y ZEA (Wam y col., 2013)
- ⇰ BEA con DON o T2 (Ruiz y col., 2011)
En algunas publicaciones se observan efectos antagónicos con presencia simultánea de tres o más micotoxinas, por ejemplo, DON, NIV y FB1; NIV, ZEA y FB1 y DON, NIV, ZEA y FB1 (Wam y col., 2013).
Yang y col. (2017) demostraron un efecto antagónico entre algunas toxinas modificadas de DON (derivados acetilados) como 15-ADON + NIV y 15-ADON + FX.
Interacciones entre micotoxinas – In vitro vs in vivo
La mayoría de estos trabajos se han efectuado in vitro, siendo la viabilidad celular el parámetro más empleado, aunque también se usan otros criterios como:
- ⇰ Apoptosis o necrosis celular
- ⇰ Daño en el ADN
- ⇰ Daño oxidativo
- ⇰ Inmunotoxicidad

Es evidente que los efectos tóxicos combinados que se observen dependen del diseño experimental:
- Tipo de células expuestas
- Tiempo de exposición
- Dosis y relación entre micotoxinas
- Puntos finales y test empleados
- Aspectos estadísticos de los modelos
Un ejemplo evidente que pone de manifiesto la importancia del diseño experimental es el de Klaric y col. (2012) que estudiaron la interacción entre OTA y citrinina (CIT) en un modelo con células epiteliales PK15 de riñón porcino, determinando el punto final mediante viabilidad celular, apoptosis, necrosis y genotoxicidad con los siguientes resultados:
- ⇰ Efecto sinérgico con respecto a viabilidad celular, apoptosis y necrosis
- ⇰ Efecto antagónico para la genotoxicidad.
Otro factor que hasta ahora ha complicado las interpretaciones es que la respuesta observada in vivo no siempre coherente con la que se da in vitro.
Para ilustrar fenómeno, podemos referirnos al ejemplo de la interacción entre DON y T-2.
En estudios in vitro, se observa antagonismo (EFSA, 2002), probablemente debido a la competencia entre ambas por los lugares de unión.
Sin embargo, los resultados in vivo con ratones (Schiefer y col., 1986) demuestran que los efectos negativos del DON en los animales se potencian con la presencia de T-2, mientras que estudios in vivo con cerdos (Friend y col., 1992) indican que DON combinada con T-2 presenta un antagonismo (con T-2 a mitad de dosis que DON).
Existen otros ejemplos publicados, como la interacción entre DON y ZEA, en los que Swamy y col. (2002) ponen de manifiesto su sinergia en lechones in vivo, mientras que Ji y col. (2017) en ratones ponen de manifiesto su antagonismo.
En cuanto a la pérdida de crecimiento observada a veces in vivo como respuesta a la presencia de micotoxinas, Andretta y col. (2015) señalan que tendría que ver con un aumento de las necesidades energéticas de mantenimiento de los animales, de acuerdo con Pastorelli y col. (2012).
En definitiva, está claro que los efectos de la combinación de micotoxinas no se pueden predecir por los efectos individuales y que, además de aditividad o sinergias, puede haber antagonismos.
Es muy difícil predecir estas respuestas al ser dosis, especie y toxina dependientes, además de la variabilidad que introducen los factores ligados a la metodología.
CONCLUSIONES
Es evidente que se debe estandarizar la metodología in vitro a nivel internacional para avanzar en el conocimiento de las interacciones entre micotoxinas y disponer de datos comparables.
Estos estándares solo serían válidos en la medida que pudieran predecir la respuesta in vivo de los animales.
El análisis de una única micotoxina puede ser insuficiente para explicar muchos casos. Sin embargo, un exceso de información analítica, sin que pueda ser correctamente interpretada, tampoco es una solución, por ello son necesarios más estudios.
En estas circunstancias, sin renunciar a otras herramientas, la mejor manera de disminuir el riesgo es la prevención a través de las buenas prácticas agrícolas y el análisis de peligros en los primeros eslabones de la cadena alimentaria.
REFERENCIAS
Almeida I, Martins HM, Santos S, Costa JM y Bernardo F. (2011). Co-occurrence of mycotoxins in swine feed produced in Portugal. Mycotoxin Research, 27, 177–181.
Andretta I, Kipper M, Hauschild L, Lehnen CR, Remus A y Melchior R. (2015). Meta-analysis of individual and combined effects of mycotoxins on growing pigs. Scientia Agricola, 73(4), 328-331.
Bensassi F, Gallerne C, el Dein OS, Hajlaoui MR, Lemaire C. y Bacha H. (2014). In vitro investigation of toxicological interactions between the fusariotoxins deoxynivalenol and zearalenone. Toxicon, 84, 1-6.
Bernhoft A, Keblys M, Morrison E, Larsen HJS y Flaoyen A. (2004). Combined effects of selected Penicillium mycotoxins on in vitro proliferation of porcine lymphocytes. Mycopathologia, 158, 441-450.
Berthiller F, Crews C, Dall’Asta C, Saeger SD, Haesaert G, Karlovsky P, Oswald IP, Seefelder W, Speijers G y Stroka J. (2013). Masked mycotoxins: A review. Molecular Nutrition & Food Research, 57, 165–186.
Biomin. The Global Mycotoxin Threat 2017.
https://info.biomin.net/acton/fs/blocks/showLandingPage/a/14109/p/p-004e/t/page/fm/17. Accedido 13/06/2019
Blajet-Kosicka A, Twaruzek M, Kosicki R, Sibiorowska E y Grajewski J. (2014). Co-occurrence and evaluation of mycotoxins in organic and conventional rye grain and products. Food Control, 38, 61-66
Driehuis F, Spanjer MC, Scholten JM, y Giffel, MC. (2008) Occurrence of Mycotoxins in Feedstuffs of Dairy Cows and Estimation of Total Dietary Intakes. Journal of Dairy Science, 91, 4261–4271.
EFSA (2002). Opinion of the Scientific Committee on Food on Fusarium toxins. Part 6: Group evaluation of T-2 toxin, HT-2 toxin, nivalenol and deoxynivalenol. SCF/CS/CNTM/MYC/27 Final.
EFSA (2011), Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA Panel on Contaminants in the Food Chain (CONTAM). EFSA Journal, 9 (10),2407.
EFSA (2014a). Scientific Opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed1 EFSA Panel on Contaminants in the Food Chain (CONTAM). EFSA Journal, 12 (12), 3916.
EFSA (2014b). Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA Panel on Contaminants in the Food Chain (CONTAM). EFSA Journal, 12 (8),3802.
EFSA (2018). Risks to human and animal health related to the presence of moniliformin in food and feed. EFSA Panel on Contaminants in the Food Chain (CONTAM), EFSA Journal, 16 (3), 5082.
Ficheux AS, Sibiril Y y Parent-Massin D. (2012). Co-exposure of Fusarium mycotoxins: in vitro myelotoxicity assessment on human hematopoietic progenitors. Toxicon, 60, 1171-1179.
Friend DW, Thompson B K, Trenholm HL, Boermans HJ, Hartin, KE y Panich PL (1992). Toxicity of T-2 toxin and its interaction with deoxynivalenol when fed to young pigs. Canadian Journal of Animal Science, 72,703–711.
Gareis M, Bauer J, Thiem J, Plank G, Grabley S y Gedek B. (1990) Cleavage of Zearalenone-Glycoside, a “Masked” Mycotoxin, during Digestion in Swine. Journal of Veterinary Medicine, 37,236–240.
Goertz A, Zuehlke S, Spiteller M, Steiner U, Dehne HW, Waalwijk C, Vries I y Oerke EC. (2010). Fusarium species and mycotoxin profiles on commercial maize hybrids in Germany. European Journal of Plant Pathology, 128, 101–111.
Grenier B, Loureiro-Bracarense A P, Schwartz H, Cossalter AM, Schatzmayr G, Moll WD y Oswald IP. (2010). Hydrolysis of Fumonisin B1 strongly reduced toxicity for piglets at the intestinal and systemic levels. 6th World Mycotoxin Forum. November 8-10. Noordwijkerhout, Netherlands.
Grenier B y Oswald IP.(2011). Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin Journal, 4, 285–313.
Gruber-Dorninger C, Novak B, Nagl V y Berthiller F. (2017). Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. Journal of Agricultural and Food Chemistry, 65, 7052−7070.
Ibáñez-Vea M, González-Peñas E, Lizarraga E y López de Cerain A. (2012). Co-occurrence of mycotoxins in Spanish barley: A statistical overview. Food Control, 28, 295–298.
Ji J, Zhu P, Cui P, Fuwei P, Zhang Y, Li Y, Wang J y Sun X. (2017). The Antagonistic Effect of Mycotoxins Deoxynivalenol and Zearalenone on Metabolic Profiling in Serum and Liver of Mice. Toxins, 9, 28.
Raj J, Farkaš H, Čepela R, Pol I, Bošnjak-Neumüller J and Vasiljević M (2018) A survey on mycotoxins detected in corn samples received from Serbia and Bosnia & Herzegovina during August to November 2017. The World Mycotoxins Forum 10th Conference, March 11-14, 2018, Amsterdam, The Netherlands (Poster no 57). Page: 116). Raj J, Farkaš H, Cepela R, Pol I, Bošnjak-Neumüller J, and Vasiljević M (2019). High level of Fumonisin B1 detected in corn samples received from Serbia during August to November 2018, 22 nd European Symposium on Poultry Nutrition (ESPN 2019), June 10-13, 2019 at Gdansk, Poland Proceedings pg 287.
Klaric MS, Zeljezic D, Rumora L, Peraica M, Pepeljnjak S y Domijan AM. (2012). A potential role of calcium in apoptosis and aberrant chromatin forms in porcine kidney PK15 cells induced by individual and combined ochratoxin A and citrinin. Archives Toxicology, 86, 97-107.
Klaassen CD y Eaton CL. (1991). Principles of Toxicology in: Toxicology, The Basic Science of Poisons. M. O. Amdur, J. Doull, and C. D. Klaassen, ed. Pergamon Press, Inc., Maxwell House, Fairview Park, Elmsford, NY, 12–49
Koshinsky HA y Khachatourians GG. (1992). Bioassay for deoxynivalenol based on the interaction of T2-toxin with trichothecene mycotoxins. Bulletin of Environmental Contamination and Toxicology, 49 (2), 246-51.
Labuda R, Parich A, Vekiru E, y Tancinová D. (2005a). Incidence of Fumonisins, Moniliformin and Fusarium species in Poultry Feed from Slovakia. Annals of Agricultural and Environmental Medicine, 12, 81–86.
Labuda R, Parich A, Berthiller F y Tančinová D. (2005b). Incidence of trichothecenes and zearalenone in poultry feed mixtures from Slovakia. The International Journal of Food Microbiology, 105, 19–25.
Marín Garcia, P. (2010). Tesis. Análisis de factores ecofisiológicos que influyen en la expresión de genes relacionados con la biosíntesis de toxinas en especies de Fusarium. Facultad de Farmacia. UNIVERSIDAD COMPLUTENSE DE MADRID.
Monbaliu S, van Poucke C, Detavernier CL, Dumoulin FdR, van De Velde M, Schoeters E, van Dyck S, Averkieva O, van Peteghem C y de Saeger S. (2010) Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin LC-MS/MS method. Journal of Agricultural and Food Chemistry, 58, 66–71.
Panasiuk L, Jedziniak P, Pietruszka K, Piatkowska M y Bocian L. (2019). Frequency and levels of regulated and emerging mycotoxins in silage in Poland. Mycotoxin Research, 35,17–25.
Pastorelli H, Milgen J, Lovatto P y Montagne L. (2012). Meta-analysis of feed intake and growth responses of growing pigs after a sanitary challenge. Animal, 6, 952-961.
Pedrosa K. (2010). Synergistic effectc of mycotoxin contaminated feed. International Pig Topics, 25 (7), 7-9.
Ruiz MJ, Franzova P, Garcia AJ y Font G. (2011). Toxicological interactions between the mycotoxins beauvericin, deoxynivalenol and T-2 toxin in CHO-K1 cells in vitro. Toxicon, 58, 315-326.
Rychlik M, Humpf HU, Marko D, Dänicke S, Mally A, Berthiller F, Klaffke H y Lorenz N. (2014). Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Research, 30, 197−205.
Schiefer HB, Hancock DS y Bhatti AR. (1986) Systemic effects of topically applied trichothecenes. I. Comparative study of various trichothecenes in mice. Journal of Veterinary Medicine, 33a, 373- 383.
Speijers GJA y Speijers MHM. (2004). Combined toxic effects of mycotoxins. Toxicology Letters, 153, 91-98.
Stoev SD, Denev S, Dutton M, Njobeh P, Mosonik J, Steenkamp P y Petkov I. (2010). Complex etiology and pathology of mycotoxic nephropathy in South African pigs. Mycotoxin Research. 26,31-46.
Streit E, Naehrer K, Rodrigues I y Schatzmayr G. (2013a). Mycotoxin occurrence in feed and feed raw materials worldwide: long-term analysis with special focus on Europe and Asia. Journal of the Science of Food and Agriculture, 93, 2892-2899.
Streit E, Schwab C, Sulyok M, Naehrer K y Schatzmayr G. (2013b). Multi-Mycotoxin Screening Reveals the Occurrence of 139 Different Secondary Metabolites in Feed and Feed Ingredients. Toxins, 5, 504-523.
Swamy HV, Smith TK, MacDonald EJ, Boermans HJ y Squires EJ, (2002b). Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent. Journal of Animal Science, 80, 3257-3267.
Thuvander A, Wikman C, y Gadhasson I. (1999). In vitro exposure of human lymphocytes to trichothecenes: Individual variation in sensitivity and effects of combined exposure on lymphocyte function. Food and Chemical Toxicology, 37, 639-648.
Trenholm HL, Cochrane WP, Cohen H, Elliot JI, Farnworth ER, Friend DW, Hamilton RM, Standish JF y Thompson BK. (1983). Survey of vomitoxin contamination of 1980 Ontario white winter wheat crop: results of survey and feeding trials. Journal Association of Official Analytical Chemists, 66 (1), 92-97.
Trenholm HL, Foster BC, Charmley LL, Thompson BK, Hartin KE, Coopock RW y Albassam MA. (1994). Effects of feeding diets containing Fusarium (naturally) contaminated wheat or pure deoxynivalenol (DON) in growing pigs. Canadian. Journal of Animal Science, 74, 361–369.
Wan LY, Turner PC y El-Nezami H. (2013a). Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food and Chemical Toxicology, 57, 276-283.
Yang Y, Yu S, Tan Y, Liu N y Wu A. (2017). Individual and Combined Cytotoxic Effects of Co-Occurring Deoxynivalenol Family Mycotoxins on Human Gastric Epithelial Cells. Toxins, 9, 96.
 Prevención de micotoxicosis
Prevención de micotoxicosis