


N. Deepa and M.Y. Sreenivasa
Molecular mycotoxicology Laboratory, Department of Studies in Microbiology, University of Mysore, Manasagangotri, Mysore, Karnataka, India.
Correspondence: +91-821-2419733; [email protected]; [email protected]
Fusarial toxins are secondary metabolites produced by Fusarium species. Fumonisins, trichothecenes, and zearalenone being the most significant ones:

Fumonisins Nearly 28 fumonisins have been identified and classified into four major groups (A, B, C, and P) among which group B is renowned as a major contaminant in food and feed. Fumonisin B (FB) is further classified into FB1, FB2, FB3 and FB4. Trichothecenes Trichothecenes are a group of secondary sesquiterpenoid metabolites produced by Fusarium species, such as F. oxysporum, F. graminearum, F. poae, F. solani, F. equisiti, F. sporotrichoides and F. xyrophilum. Based on the functional group placement, trichothecenes are further classified into type A, B, C and D, among which type D is the most toxic one and type A and B have mixed toxicity Type A trichothecenes –T-2 toxin (T-2) and HT-2 toxin (HT-2)- and type B trichothecenes –nivalenol (NIV) and deoxynivalenol (DON)– are the most relevant ones in this group (Zhang et al., 2020). Zearalenone Zearalenone is produced by F. culmorum, F. cerealis, F. semitectum, F. equiseti and F. graminearum (Krol et al., 2018).![]() FB1 represents 70% of the reported fumonisins with a prominent distribution worldwide (Deepa y Sreenivasa et al., 2017a, 2017b).
FB1 represents 70% of the reported fumonisins with a prominent distribution worldwide (Deepa y Sreenivasa et al., 2017a, 2017b).
Distribution of fusarial toxins
Fusarial toxins can be found in food, feed, freshly harvested and stored agricultural commodities worldwide.![]() They are highly associated with corn and corn-based products, but also can be found in paddy, sorghum, pearl millet, wheat, beans, curry, barley, asparagus, chilli pickle, cowpea, triticale, soybeans, red wine, beer, onion, garlic, black radish, black tea, coffee, grapes, cassava products, figs, peanuts, milk, meat, egg, leaves of orange, dietary plants, mint, sage leaves, medicinal plants, brewing adjuncts, valerian root, linden flowers and chamomile in China, Tanzania, Ivoire, Italy, South Africa, Portugal, India, Turkey and Spain (Ma et al., 2018) (Table 1).
They are highly associated with corn and corn-based products, but also can be found in paddy, sorghum, pearl millet, wheat, beans, curry, barley, asparagus, chilli pickle, cowpea, triticale, soybeans, red wine, beer, onion, garlic, black radish, black tea, coffee, grapes, cassava products, figs, peanuts, milk, meat, egg, leaves of orange, dietary plants, mint, sage leaves, medicinal plants, brewing adjuncts, valerian root, linden flowers and chamomile in China, Tanzania, Ivoire, Italy, South Africa, Portugal, India, Turkey and Spain (Ma et al., 2018) (Table 1).
Table 1. Fusarium species associated with food and feed and their mycotoxins.
Mechanisms of action and degradation of fusarial toxins Fumonisins
Fumonisins inhibit the function of the key enzyme ceramide synthase, interrupting the completion of sphingolipid metabolism in cells, hepatocytes, tissues, renal cells, and neurons. Biosynthesis of sphingolipids occurs in the endoplasmic reticulum through a process of hydrolysis from complex sphingolipids into ceramides and, later, to sphingosine, which is phosphorylated and cleaved to fatty aldehyde and ethanolamine phosphate for further incorporation into phosphatidyl-ethanolamine. Alteration of sphingolipid biosynthesis can play a crucial role in carcinogenesis and disease development by damaging DNA (Alizadeh et al., 2012) (Figure 1).
Figure 1. Mechanism of action of Fumonisin B1. Trichothecenes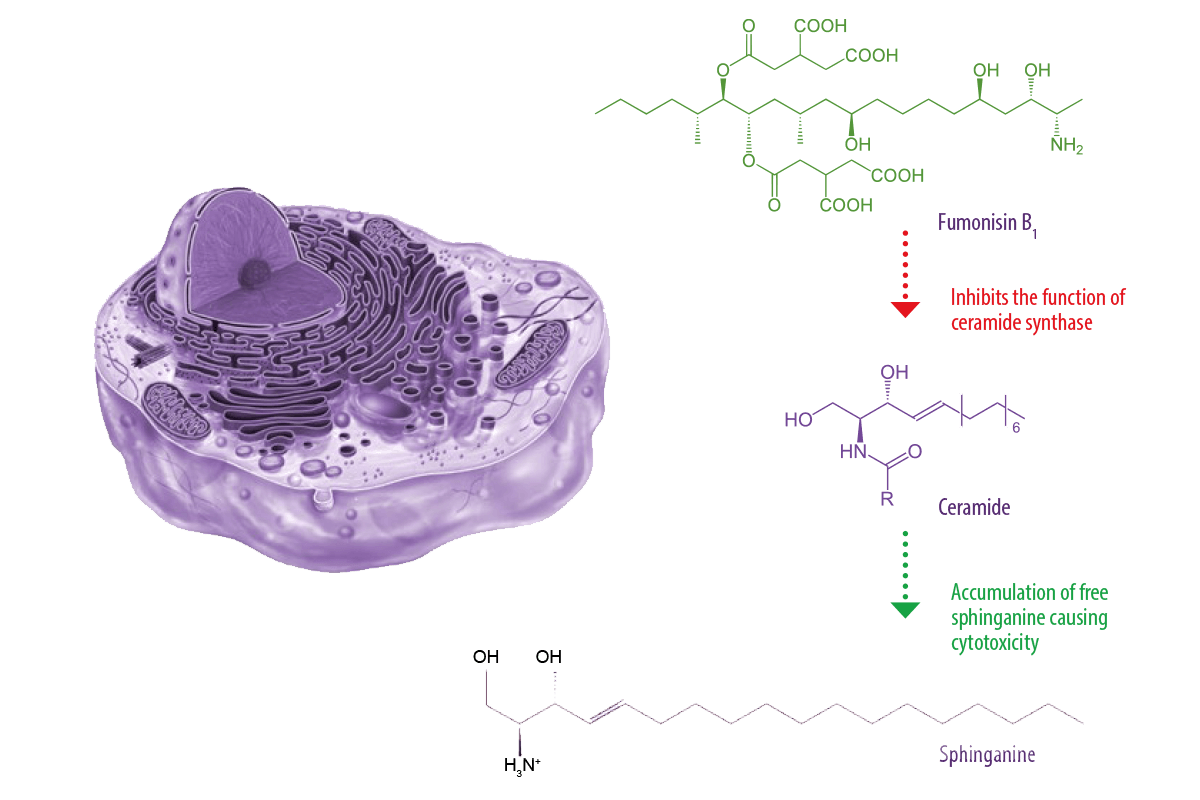
Trichothecene degradation is mainly based on their epoxide group and acylated side chains (acylated trichothecenes referred to T-2 toxin and non-acylated trichothecene referred to DON). Degradation of trichothecenes involves two mechanisms:
![]() Some reports state that oxidation or isomerization, hydroxylation and glycosylation are also pathways involved in trichothecene degradation, forming compounds that can later be regenerated or rehydrolyzed in the digestive tract of humans and animals (Wang et al., 2020).
Some reports state that oxidation or isomerization, hydroxylation and glycosylation are also pathways involved in trichothecene degradation, forming compounds that can later be regenerated or rehydrolyzed in the digestive tract of humans and animals (Wang et al., 2020).
Zearalenone ZEN metabolism involves two degradation mechanisms: 1. Cleaving a lactone ring structure into a less 2. Another pathway occurs through a lactone intermediate without decarboxylation, observation of estrogenic activity (Tintelnot et al., 2011).
toxic compound (α-zearalanol and β-zearalenol). The enzyme ZEN lactonohydrolase supports this pathway, resulting in damaging estrogenic activity through decarboxylation (Popiel et al., 2014).
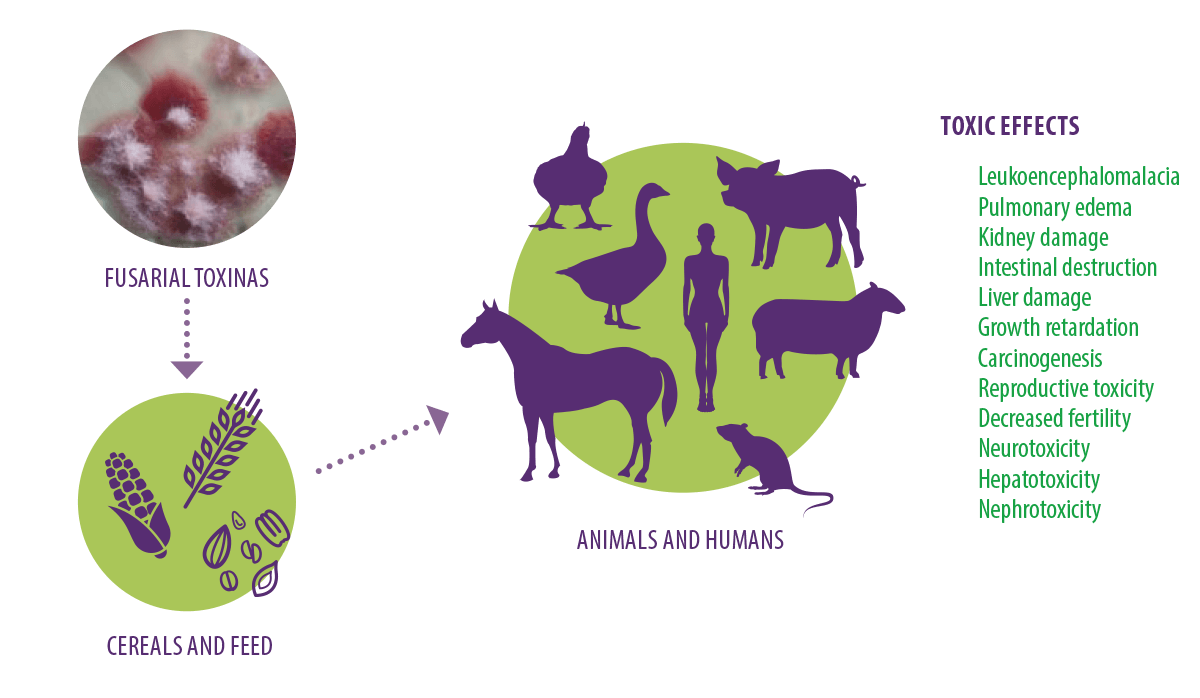
Effects of fusarial toxins Fusarial toxins have important toxicological effects in humans and animals. They have proven to be nephrotoxic, carcinogenic, hepatotoxic, hepatocarcinogenic and cytotoxic in mammalian cells, with estrogenic, immunotoxic, and emetic effects, leading to intestinal, lung, liver, and kidney damage (Yu et al., 2021) (Figure 2). Furthermore, ingestion of fusarial toxin-contaminated feed is linked to leukoencephalomalacia in horses, and pulmonary edema syndrome and hydrothorax in pigs (Kaminski et al., 2020). Figure 2. Toxic effects of fusarial mycotoxins. Fumonisins
FB1 derived sphingolipids display intricate protagonism in many cell function processes. Accumulation of sphingoid bases causes cell growth inhibition, leading to cytotoxicity and interfering with protein kinase C, Na+/K+ ATPase, triggering or constraining enzymes involved in lipid signaling pathways and activating phospholipase D, as well as interfering with dephosphorylation of retinoblastoma protein. ⇰ This process increases the risk of cancer via lipid mediators and apoptosis that control cell proliferation. Ceramide synthase inhibition leads to the accumulation of free sphinganine in the kidneys, liver, and lungs. ⇰ This hydrophobic compound is capable of crossing cell membranes and shows up in urine and blood. This mechanism targets the kidney and liver in most animals by increasing oncotic necrosis, regeneration, apoptotic, and bile duct hyperplasia (Liu et al., 2019). Fumonisins are also known to be phytotoxic. ⇰ Studies have shown that FB1 is harmful for pigweed, hemp sesbania, and duckweed, causing chlorophyll loss and stunted growth.![]() Therefore, FB1 is considered a Group 2B carcinogen in humans by th International Agency for Research on Cancer (IARC) since 2002.
Therefore, FB1 is considered a Group 2B carcinogen in humans by th International Agency for Research on Cancer (IARC) since 2002.
Zearalenone ZEN has cytogenetic toxicity, leading to decreased fertility as well as anti-androgenic effects in pigs, cattle, and sheep (Feizollhi et al., 2020). Trichothecenes The 12,13-epoxy-trichothec-9-ene nucleus found in trichothecenes is necessary for its toxicity, inhibiting DNA and RNA synthesis, affecting intestinal integrity, and causing immunotoxicity and emetic effects.
Fusarial toxin remediation Due to the toxicity of fusarial toxins, it is important to implement measures to reduce contamination in food and feed matrixes to decrease the risk of health hazards among humans and animals (Buszewska-Forajta et al., 2020). International organizations such as the European Food Safety (EFSA), World Health Organization (WHO) and Food and Agriculture Organization (FAO) have set up strict control measures, establishing maximum residue levels in food and feedstuffs, for example: In this context, many remediation strategies have been put in place, including physical, chemical, and biological methods (Li et al., 2021). Biodegradation of fusarial toxins Biological detoxification methods are efficient, specific, and environment-friendly. Although the microbes used in them have the limiting factor of requiring certain growth conditions, due to their enormous beneficial applications for toxin degradation, biodegradation of fusarial toxins with bio-adsorbents and enzymes is a promising strategy that has attracted the attention of the scientific community (Deepa et al., 2021) (Figure 3). Figure 3. Biodegradation of fusarial toxins. Toxic groups in fumonisins, such as tricarboxylic acid group (-TCA) and amino group (-NH2), inhibit the sphingosine N-acetyltransferase during sphingolipid metabolism, blocking the sphingolipid signaling and affecting cell differentiation, apoptosis, and proliferation. Fumonisin biodegradation It has been shown that the presence of FB1 in intestinal cells inhibits the ceramide synthase enzyme, forming acetylated N-acyl HFB1, which is more toxic than HFB1, reducing body weight and disrupting the gut microbial balance in broilers (Yu et al., 2022).


⇰ Evaluated in GES-1 cells, it proved to be safe for controlling FB contamination in feed and food (Li et al., 2022).![]() In a study, a fusion enzyme was designed for the degradation of FB1 using carboxylesterase and aminotransferase. This enzyme was cloned to vector pPIC9K, successfully expressed in the host Pichia pastoris GS115, and named FUMD1.
In a study, a fusion enzyme was designed for the degradation of FB1 using carboxylesterase and aminotransferase. This enzyme was cloned to vector pPIC9K, successfully expressed in the host Pichia pastoris GS115, and named FUMD1.![]() Saccharomyces cerevisiae strain IS1/1 and SC82 degraded FB1 by 45% and 22%, and a mixture of FB1 and FB2 by 50% and 25%, respectively.
Saccharomyces cerevisiae strain IS1/1 and SC82 degraded FB1 by 45% and 22%, and a mixture of FB1 and FB2 by 50% and 25%, respectively.![]() Camilo et al. (2000) applied three strains of Bacillus species (S9, S10, S11) that degraded FB1 by 43%, 48%, and 83%, respectively.
Camilo et al. (2000) applied three strains of Bacillus species (S9, S10, S11) that degraded FB1 by 43%, 48%, and 83%, respectively.![]() Complete degradation of FB1 was observed 24h after applying the strain NCB 1492 and bacterial consortium SAAS79 (Benedetti et al., 2006).
Complete degradation of FB1 was observed 24h after applying the strain NCB 1492 and bacterial consortium SAAS79 (Benedetti et al., 2006).![]() FumD enzyme completely degraded FB1 to HFB1 in the duodenum and jejunum of pigs in turkeys (Schaumerger et al., 2016).
FumD enzyme completely degraded FB1 to HFB1 in the duodenum and jejunum of pigs in turkeys (Schaumerger et al., 2016).![]() The black yeast Exophiala spinifera degraded FB1 to AP1 (Polyolamine) and 2-OP1 by extracellular enzyme carboxylesterase.
The black yeast Exophiala spinifera degraded FB1 to AP1 (Polyolamine) and 2-OP1 by extracellular enzyme carboxylesterase.![]() Recently, Azotobacter was shown to be a key bacterium for degradation of FB1, reporting 98% of degradation after a 2-hour incubation (Deepa et al., 2022).
Recently, Azotobacter was shown to be a key bacterium for degradation of FB1, reporting 98% of degradation after a 2-hour incubation (Deepa et al., 2022).![]() Deepthi et al. (2016) reported FB1 degradation by 61.7% after treating with Lactobacillus plantarum MYS6.
Deepthi et al. (2016) reported FB1 degradation by 61.7% after treating with Lactobacillus plantarum MYS6.
ZEN, DON, DAS, and T-2 toxin biodegradation
⇰ However, certain reports suggest that ZEN-derived products are more toxic than the ZEN (zearalenol > α-zearalanol > zearalenone > β-zearalenol) (Krol et al., 2018). ![]() Some lactic acid bacteria, such as Pseudomonas otitidis (Tan et al., 2015), Rhodococcus pyridinivorans strains K408/ AK37 (Cserhati et al., 2013), and Bacillus velezensis strain ANSB01E (Guo et al., 2019) degrade and detoxify ZEN and T-2 toxin simultaneously (Barlkiene et al., 2018).
Some lactic acid bacteria, such as Pseudomonas otitidis (Tan et al., 2015), Rhodococcus pyridinivorans strains K408/ AK37 (Cserhati et al., 2013), and Bacillus velezensis strain ANSB01E (Guo et al., 2019) degrade and detoxify ZEN and T-2 toxin simultaneously (Barlkiene et al., 2018).![]() Saccharomyces cerevisiae CECT 1891 and L. acidophilus 24 act as an adsorbent, degrading ZEN, DON, and FB1 (Campaginolloo et al., 2015).
Saccharomyces cerevisiae CECT 1891 and L. acidophilus 24 act as an adsorbent, degrading ZEN, DON, and FB1 (Campaginolloo et al., 2015).![]() Ery4 laccase enzyme from Pleurotus eryngii degrades multiple mycotoxins, including FB1, ZEN and T-2 toxin (Lio et al., 2018).
Ery4 laccase enzyme from Pleurotus eryngii degrades multiple mycotoxins, including FB1, ZEN and T-2 toxin (Lio et al., 2018).![]() Microbiota isolated from chicken intestines had the capability to degrade nearly 12 trichothecenes through de-epoxidation and diacylation (Young et al., 2007).
Microbiota isolated from chicken intestines had the capability to degrade nearly 12 trichothecenes through de-epoxidation and diacylation (Young et al., 2007).![]() Rat microbiota, through de-epoxidation, transformed T-2 toxin into HT-2 toxin, as well as de-epoxy T-2 triol and DAS to de-epoxyscirpentriol and de-epoxymoniacetoxyscirpenol.
Rat microbiota, through de-epoxidation, transformed T-2 toxin into HT-2 toxin, as well as de-epoxy T-2 triol and DAS to de-epoxyscirpentriol and de-epoxymoniacetoxyscirpenol.![]() Pig gut microbiota degraded ZEN through hydrolysis into α-zearalenol and DON into de-epoxy DON through de-epoxydation mechanism.
Pig gut microbiota degraded ZEN through hydrolysis into α-zearalenol and DON into de-epoxy DON through de-epoxydation mechanism.![]() Eggerthella species DII-9 degrade up to 86% trichothecenes (DON, T2 triol, HT-2, T-2 tetraol) through de-epoxidation (Gao et al., 2018).
Eggerthella species DII-9 degrade up to 86% trichothecenes (DON, T2 triol, HT-2, T-2 tetraol) through de-epoxidation (Gao et al., 2018).![]() Bacillus pumilus ES-21 degraded ZEN more than 95.7% with release of degraded product 1-(3,5-dihydroxyphenyl)-60- hydroxy-l0-undecen-l00-one through esterase activity (Wang et al., 2017).
Bacillus pumilus ES-21 degraded ZEN more than 95.7% with release of degraded product 1-(3,5-dihydroxyphenyl)-60- hydroxy-l0-undecen-l00-one through esterase activity (Wang et al., 2017).![]() Bacillus amyloliquifaciens degraded ZEN through extracellular enzymes without any derivatives (Xu et al., 2016).
Bacillus amyloliquifaciens degraded ZEN through extracellular enzymes without any derivatives (Xu et al., 2016). ![]() Nine biocontrol agents from Aspergillus y Rhizopus detoxified ZEN, resulting in five degraded compounds α- zearalenol-sulfate, ZEN- O-16- glucoside, α-zearalenol, ZEN-14-sulfate and ZEN-O-14 (Koch et al., 2014).
Nine biocontrol agents from Aspergillus y Rhizopus detoxified ZEN, resulting in five degraded compounds α- zearalenol-sulfate, ZEN- O-16- glucoside, α-zearalenol, ZEN-14-sulfate and ZEN-O-14 (Koch et al., 2014). ![]() Bifidobacterium and Lactococcus lactis from milk neutralized ZEN by absorption process up to 88% (Mokoena et al., 2005).
Bifidobacterium and Lactococcus lactis from milk neutralized ZEN by absorption process up to 88% (Mokoena et al., 2005).
CONCLUSIONS Food and feed contamination with fusarial toxins poses a serious risk to human and animal health, not to mention the economic costs they entail for farmers. Biodegradation plays a dynamic role as a reliable, efficient, simple, and environmental-friendly method that is less costly in comparison to other methods. However, there is still little information available on the mechanisms involved in the degradation of fusarial toxins.![]() Therefore, it is necessary to conduct further research to shed light on these processes and their possible applications in the field.
Therefore, it is necessary to conduct further research to shed light on these processes and their possible applications in the field.
 Micotoxicosis prevention
Micotoxicosis prevention